De evolutietheorie ontkracht
Charles Darwin had het mis
Fossiel bewijs voor evolutie van de amfibieën
Voor de evolutie van vissen naar vierpotige amfibieën zou overweldigend veel bewijsmateriaal bestaan. Er zouden talloze overgangsvormen zijn gevonden, waaronder het bekende Tiktaalik fossiel. Dit zou het bewijs vormen dat alle viervoetige landdieren, en dus ook wij, afstammen van de vissen.

Laten we nu even naar de vinnen en poten kijken van deze zogenaamde overgangsvormen:
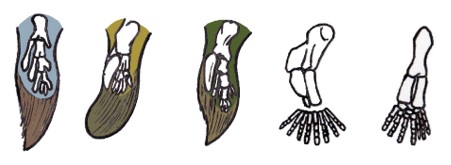
We zien dat deze zogenaamde overgangsvormen toch wel vrij problematisch zijn en in geen geval een ‘mooie overgang’ tonen. Tiktaalik zou de gevonden ‘missing link’ zijn tussen de vis Panderichtys en het amfibie Acanthostega. Echter, de vin (met vinstralen!) van Tiktaalik lijkt meer op die van Eusthenopteron dan op die van zijn ‘directe voorouder’. Het is ook belangrijk om op te merken dat deze afbeeldingen niet in werkelijke verhouding t.o.v. elkaar zijn getekend: die vinnen en poten zijn in geen geval even groot, want die soorten waren in geen geval even groot. Eusthenopteron was 1,5 tot 1,8 meter[1] en werd gevonden in Quebec, Canada (Noord-Amerika) Panderichtys was 0,9 tot 1,3 meter[2] en werd gevonden in Letland (Europa); Tiktaalik was ruim 3 meter en werd dan weer in Canada gevonden; Acanthostega, die uit Tiktaalik zou zijn geëvolueerd, was slechts 60 centimeter en werd gevonden in Groenland.[3]; Ichtyostega die uit Acanthostega zou zijn geëvolueerd, was dan weer 1,5 meter en werd ook in Groenland gevonden. De allerlaatste in de rij is Pederpes, die 1 meter groot werd en in Schotland werd gevonden.[4] Het evolutieschema ziet er dan niet uit zoals op de vorige bladzijde, maar zo:

Dat begint al problematisch te worden. Aan de Gentse universiteit toonden ze ons het eerste diagram. Het begint duidelijk te worden dat het verhaal over de evolutie van de amfibieën langs alle kanten begint te rammelen. Laten we nu eerst even Eusthenopteron bekijken. Dit geslacht van vissen werd ondergebracht in de orde van de kwastvinnigen (Sarcopterygii) waartoe ook de nog bestaande coelacanten (Latimeria) en longvissen (Dipnoi) behoren. Maar ook de Tetrapoda, en dus alle landdieren inclusief de mens, wordt volgens de cladistische (en dus evolutionistische) indeling, of het evolutie-cladogram, in deze klasse ondergebracht, omdat wij zouden afstammen van deze vissen. Het is inderdaad wel zo dat Eusthenopteron gelijkenissen vertoont met de coelacant, die, zoals we reeds gezien hebben, er identiek hetzelfde uitziet als miljoenen jaren oude fossielen.

Zowel Eusthenopteron als de coelacant (Latimeria) heeft 8 vinnen (staartvin niet meegerekend). De grootte van de vissen is eveneens quasi identiek: 1,5 tot 1,8 meter. De kwastvinnen werden verondersteld primitieve poten te zijn. Eusthenopteron wordt dan ook vaak al wandelend, half uit het water of zelfs op het droge afgebeeld. Ondertussen is men grotendeels van deze piste afgestapt, en wordt Eusthenopteron als volledig aquatisch beschouwd. Het feit dat ‘wandelen op de zeebodem’ en ‘wandelen op het droge’ bij de huidig bestaande coelacant (Latimeria), die aan de rand van het continentaal plat leeft, op diepten van 150 tot 700 meter, niet werd waargenomen, was uiteraard problematisch. Het hebben van kwastvinnen is geen bewijs dat de vis aan het evolueren is in een viervoetig landdier.


Dan heb je Tiktaalik, dé zogenaamde overgangsvorm tussen de laatste vis en het eerste amfibie in het evolutionaire schema. Deze vis wordt steevast half uit het water afgebeeld, alsof het dier op het droge kwam kruipen en aldus aan het ‘evolueren’ was in een landdier.

Er zijn natuurlijk enkele problemen. Eerst en vooral was dit dier naar schatting ruim 3 meter lang, in tegenstelling tot zijn evolutionaire ‘voorouder’, die tot 1,3 meter werd, en zijn evolutionaire ‘opvolger’ die slechts 60 centimeter werd. Deze enorme variatie in afmetingen is al problematisch. Verder heb je het probleem dat ze geen 100% compleet skelet hebben teruggevonden. De reconstructies (zoals op de figuur) zijn gedeeltelijk speculatie. Ook de veronderstelling dat deze vis op het droge kwam, neusgaten en longen had, is puur speculatie. Er is daar geen enkel fossiel bewijs voor gevonden. De kwastvinnen zijn nog steeds vinnen, en geen poten (zie ook fig. op blz. 71) en de fossiele schedel toont géén neusgaten. Ook wordt vaak de bijzondere vorm van de schedel aangewend als argument dat dit een voorouder zou zijn van de Tetrapoda. Maar er zijn heel wat vissen met bizarre schedels. Neem nu de kaaimansnoek.

Maakt het hebben van deze bijzondere schedelvorm deze vis nu tot voorouder van een bepaalde groep viervoetige landdieren die daar sterk op lijken? Ik dacht het niet. Verder heb je de longvissen, die ook kwastvinnen hebben. De meeste longvissen hebben echter zeer vreemde kwastvinnen, die in geen geval op ‘evoluerende’ poten lijken. Longvissen zijn aangepast aan extreme omstandigheden, zodat die in extreem droge perioden ook op land kan overleven, bij voorkeur in een hol in het slijk. Maar tijdens het grootste gedeelte van zijn leven, verblijft de longvis in het water, en gebruikt hij z’n kieuwen voor ademhaling. Longvissen worden dan ook niet beschouwd als de evolutionaire voorouder van de amfibieën.

Eusthenopteron, Tiktaalik en Latimeria worden allemaal cladistisch ingedeeld binnen de kwastvinnigen (Sarcopterygii). Het zouden de kwastvinnigen geweest zijn die aanleiding gaven tot de Tetrapoda, of de viervoetige landdieren. Maar er bestaan heel wat vissen binnen de groep van de straalvinnigen (die vanuit evolutionair oogpunt géén aanleiding gaven tot de Tetrapoda) die hun vinnen gebruiken om over de zeebodem te wandelen, of in nood op het droge zich voort te bewegen (zoals Periophtalmus).




Zowel sommige kwastvinnige vissen als sommige straalvinnige vissen gebruiken vinnen om zich op de bodem voort te bewegen (hetzij op de zeebodem, hetzij op het droge), terwijl andere kwastvinnige vissen en straalvinnige vissen hun vinnen gebruiken enkel om te zwemmen. Zijn die vissen die hun vinnen gebruiken dan om ‘te wandelen’ nu bewijs voor evolutie van amfibieën uit vissen? Neen, want dit fenomeen doet zich in beide groepen (kwastvinnigen en straalvinnigen) voor, en zowel bij soorten die zich op het land begeven en longen hebben; soorten die zich op het land begeven en geen longen hebben, en soorten die zich niet op het land begeven. Dus het gebruik van vinnen om te steunen, of zich ‘al wandelend’ voort te bewegen is in geen geval een overtuigend argument voor evolutie van Tetrapoda.
Verder zijn de fossielen ook problematisch, in die zin dat men niet kan weten of Tiktaalik reeds bruikbare longen had – zoals de longvissen bijvoorbeeld – of niet. Men veronderstelt van wel. Hedendaagse kwastvinnige vissen hebben volwaardige kieuwen, maar ook longen (die al dan niet gebruikt worden). Men veronderstel dat Tiktaalik een kwastvinnige vis was, die reeds uit het water en op het land kon leven – net zoals de hedendaagse longvissen. Maar zijn zogenaamde directe evolutionaire opvolger – die overigens 5 keer kleiner was – was een amfibie. En hier komt een heel groot probleem opduiken: amfibieën kennen, in tegenstelling tot vissen, metamorfose.[5]
De combinatie van de enorme verschillen in afmetingen tussen de zogenaamde overgangsvormen, het gebrek aan intermediaire vormen tussen kwastvinnen en volwaardige poten, het feit dat amfibieën metamorfose kennen en het feit dat zowel kwastvinnigen als straalvinnigen hun vinnen gebruiken om te ‘wandelen’, is uitermate problematisch voor evolutie van Tetrapoda uit kwastvinnige vissen.
En dan hebben we het nog niet eens gehad over de vermeende ouderdom van die fossielen.
[1] https://www.britannica.com/animal/Eusthenopteron
[2] http://www.prehistoric-wildlife.com/species/p/panderichthys.html
[3] http://www.prehistoric-wildlife.com/species/a/acanthostega.html
[4] http://www.prehistoric-wildlife.com/species/p/pederpes.html
[5] Zie hoofdstuk 5.2: Van larve naar adult.
[…] Maar ook bij gewervelden is er één groep die metamorfose kent: de amfibieën. Daar heb je namelijk het aquatische stadium met dikkopjes: dat zijn een soort larven zonder poten, met een staart en kieuwen (geen zijvinnen!); en het volwassen stadium met poten, longen en met staart (salamanders) of zonder staart (kikkers en padden). Alle andere gewervelden (vissen, reptielen, vogels en zoogdieren) kennen geen zo’n metamorfose tijdens de ontwikkeling: het individu dat geboren wordt (juveniel) is opnieuw een miniatuurversie van het volwassen individu (adult); er is enkel ontwikkeling.

Bij metamorfose moet de genetische informatie aanwezig zijn voor alle stadia, en dus ook voor het verlies bepaalde kenmerken zoals de kieuwen en de staart bij kikkers of de aanmaak van vleugels, een lange roltong enz. bij vlinders. Is het feit dat de larve van een aasvlieg eruit ziet als een worm nu bewijs dat de vlieg afstamt van een worm? Of het feit dat een dikkopje er wat visachtig uitziet, bewijs dat kikkers van vissen zouden afstammen? Neen. Metamorfose is een uniek kenmerk voor bepaalde diersoorten, maar het is eigenlijk geen bewijs voor evolutie. Het is zelfs eerder problematisch.
Vooral het feit dat amfibieën metamorfose kennen is eigenlijk zeer problematisch voor evolutie van amfibieën uit kwastvinnige vissen, zoals Tiktaalik, die als dé voorouder van de amfibieën, en daarmee alle gewervelde landdieren wordt beschouwd.
Kwastvinnige vissen, zoals ook Tiktaalik er één was, hebben zowel kieuwen als longen. Maar vissen ondergaan geen metamorfose. Vanaf het moment van de bevruchting, ontwikkelen ze zowel kieuwen en longen, als kwastvinnen. Nergens in het DNA van kwastvinnigen is er informatie voorzien voor metamorfose. Maar amfibieën daarentegen, hebben in het juveniel stadium géén longen, maar wel kieuwen. Gaandeweg worden de volwaardige poten ontwikkeld en verandert de staartvin in een gewone staart – of verdwijnt deze in het geval van kikkers en padden. De kieuwen verdwijnen en er komen longen in de plaats. Dit valt in geen enkel opzicht te rijmen met evolutie van amfibieën uit een kwastvinnige. Opvallend is vooral: men heeft het over de evolutie van de kwastvin in een amfibieënpoot, maar bij de metamorfose bij amfibieën is er geen sprake van vinnen die in poten veranderen – de poten ontstaan gewoon op de plaats waar ze moeten komen, zonder dat er daar vinnen waren.
